底质革命示意图,从埃迪卡拉纪(左)到寒武纪(右)。(图片来源:sepmstrata.org)
有研究提出,在所有对称性设计中,两侧对称能够为动物改变运动方向提供最有效的推动力。可以说,两侧对称的身体构造,为动物们真正“动”起来提供了基础,让地球变成了物种间频繁竞争、捕食的运动场。当然,两侧对称的适应意义不仅限于运动能力,也有研究提出,两侧对称的结构能提高动物体内物质输送的效率。
源远流长
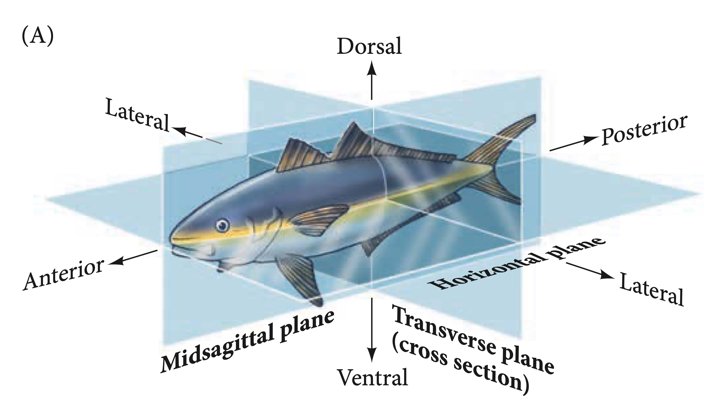
回看历史,我们两侧对称的身体源自多细胞动物的早期演化;而在每一只两侧对称动物的发育过程中,胚胎的对称性也经历了复杂的变化,受到了相关发育基因的精细调控。在发育生物学描述中,两侧对称动物在三维空间中的两个坐标轴上存在差异:区分前端和后端的前后(anterior-posterior,AP)轴,和区分背面与腹面的背腹(dorsal-ventral,DV)轴。体轴的建立与维持,是胚胎发育过程中的大事。
文章插图
AP轴与DV轴示意图。Developmental Biology. Eleventh Edition. By Scott F. Gilbert and Michael J. F. Barresi.
众所周知,胚胎发育的起点是看起来没有什么“轴”可言的一个细胞——受精卵。虽然受精卵的细胞质根据所含卵黄的多少就已经分出了植物极-动物极的极性,但前后轴和背腹轴的真正建立,始于动物界特有的胚胎发育阶段——原肠胚期。著名发育生物学家刘易斯·沃尔珀特(Lewis Wolpert)有云:“ 你一生中最重要的时刻不是出生、结婚或死亡,而是原肠胚形成。”在这一时期,胚胎卷曲折叠,内部的细胞迁移运动,形成了不同的胚层,两侧对称动物胚胎的前后轴和背腹轴基本架构也显现出来。

文章插图
两栖类胚胎的原肠化。(图片来源:www.zum.de)
说到这里,你肯定有一个疑问:胚胎的这种有规律的原肠运动,肯定不是随机的,得有个指挥信号,告诉胚胎在哪个方向上产生头尾的分化,哪个方向产生背腹的分化等等。这个信号,就是一些调控下游基因表达的转录因子蛋白。这些所谓的“形态发生素(morphogen)”在胚胎不同位置的浓度有所差异,所形成的浓度梯度,就定义了体轴。
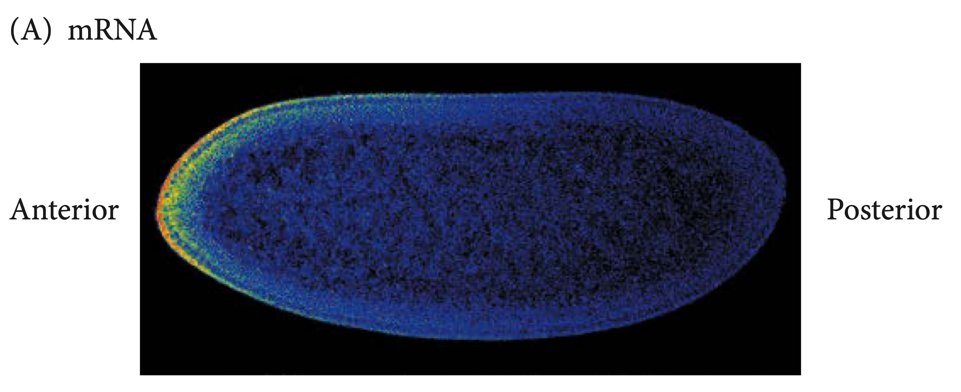
例如,在发育生物学的模式物种果蝇中,只在卵细胞一侧与卵细胞接触的滤泡细胞,会向卵细胞内分泌一种激酶,让卵细胞内的微管结构定向排列;两个基因bicoid和oskar的mRNA在微管上结合不同的转运蛋白,分别沿微管向不同方向运输;这导致bicoid mRNA在卵细胞的“前端”聚集,而在“后端”,翻译产生的Oskar蛋白结合富集了nanos基因的mRNA。当卵细胞受精并开始细胞核的分裂时,前后端分别翻译出高浓度的Bicoid和Nanos蛋白,这两种形态发生素的相反浓度梯度,就定义了原肠胚时期胚胎运动的前后轴,并引发下游调控因子的区域性表达。一波又一波的形态发生素最终将胚胎划分成多个体节;最后,大名鼎鼎的Hox基因家族的不同成员在不同体节中顺序表达,定义了头、胸、腹部各个体节迥异的结构和功能。
文章插图
bicoid mRNA在果蝇卵细胞中的分布,左侧为前端。Developmental Biology. Eleventh Edition. By Scott F. Gilbert and Michael J. F. Barresi.
在果蝇所在的双翅目类群之外,绝大多数多细胞动物的前后轴都是由Wnt基因家族从胚胎后端表达的调控因子蛋白来定义的。同样地,在绝大多数两侧对称动物中,背腹轴的确立来自于BMP基因家族的调控因子浓度梯度。有趣的是,BMP家族蛋白的高浓度表达,在我们脊椎动物中定义了中枢神经所在的背部,而在其他两侧对称动物中定义的是同样包含中枢神经的腹部。经历了五亿年分道扬镳的独立演化后,我们人类身体的最基本结构,仍然与小小的昆虫使用着同宗同源的定义方式。
- |连“拍马屁”都不懂,怎么混职场?难怪圆滑的人比老实人更受欢迎
- 新乡|求职者太多影响其它公司,质疑“占用公共资源”
- 周树人$为何鲁迅对藤野先生“永志难忘”,而藤野却记不起“鲁迅是谁”
- 招聘|“血奴”事件曝光后,更多骗局被扒出,招聘网站陷入“至暗时刻”
- |这4类教师不配当“老师”,将面临失业,家长们:早该这样了
- 保尔@张海迪与王佐良“婚礼只是形式,最重要的是一辈子都不离不弃”
- 孙悟空$俗语“朋友妻不可欺”,后面其实还有一句,更加经典,却少有人知
- 姑父#“人有三不亲,姨父、姑父”,另一个是谁?说了你别不放心上
- 发财&俗话“一代做官九代绝”,下半句更加经典,古人智慧,道尽了现实
- 鲁智深$大家都认为林冲软弱可欺,为什么金圣叹说他“太狠”?